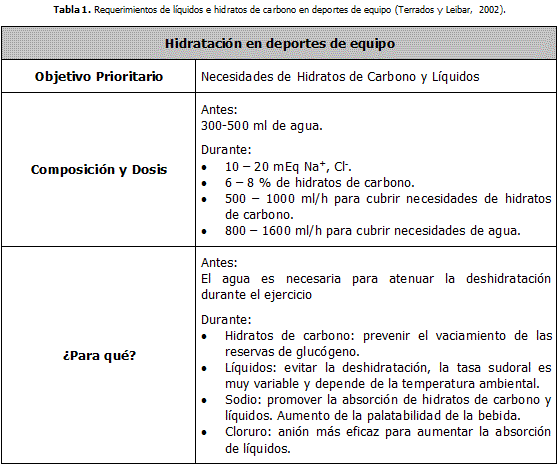
Stearoyl-CoA desaturase-1 (SCD1) es una enzima del retículo endoplasmático que contiene hierro microsomal, esta enzima es necesaria para la formación endógena de ácidos grasos monoinsaturados (MUFA) a partir de ácidos grasos saturados (SFA). SCD1 convierte SFA a MUFA por incorporación de un doble enlace entre los carbonos 9 y 10 de los ácidos grasos de cadena larga acyl-CoAs, principalmente en Stearoyl-CoA (18:0) o Palmitoyl-CoA (16:0), para generar Oleoyl-CoA (18:1n9) o Palmitoleoyl-CoA (16:1n9), respectivamente (16). La reacción de desaturación requiere NADH/NAD +, citocromo b6, citocromo b6 reductasa, y el oxígeno.
SCD1 es una enzima limitante que es esencial para la conversión de SFA endógenos y exógenos en MUFA encontrada en los mamíferos. Hay dos isoformas identificadas en humanos (SCD1 y SCD5). La isoforma SCD1 es sumamente expresada en el músculo esquelético y parece influir en la composición ácidos grasos, en la esterificación de lípidos, lipogenesis, y ßeta-oxidación, todo lo cual indirectamente influye en la respuesta de la glucosa y la señalización de insulina. Sin embargo, la mayoría de investigaciones que implica SCD1 se han enfocado mucho más en la expresión hepática SCD1 y no la expresión en el músculo.
El Índice de desaturación y la Composición ácidos grasos como Indicadores Indirectos para la Expresión SCD1 en el Músculo Esquelético
Mientras hay una escasez de datos que exclusivamente enfoca la expresión SCD1 en el músculo, se pueden hacer inferencias sobre la expresión SCD1 sobre el músculo (Tabla1), evaluando ciertos sustitutos de los marcadores actuales. Estas inferencias sobre la expresión de SCD1 incluyen el análisis de composición ácidos grasos y el índice de desaturación (DI) en entrenados por ejercicio contra grupos sedentarios y poblaciones delgadas y obesas.

La Composición del Cuerpo y el Nivel de Actividad Cambian la Expresión SCD1 en Músculo Esquelético
Diferencias han sido observadas cuando la expresión SCD1 en el músculo de grupos delgados y obesos han sido evaluadas. La expresión SCD1 ha mostrado ser elevada en el sujeto humano obeso. El aumento de la expresión SCD1 fue correlacionado con cambios de la composición ácidos grasos de glicerolípidos con aumento oleico y disminución del ácido palmítico (16:0) y el ácido esteárico (18:0) en sujetos flacos. DI (16:1/16:0 y 18:1/18:0) era también más alto en el grupo de obesos comparado con el grupo de delgados. De modo interesante, la expresión aumentada de SCD1 en sujetos obesos estaba correlacionada con disminución de la ßeta-oxidación.
Un sólo de ejercicio aeróbico ha sido demostrado que es capaz de aumentar las concentraciones de proteína del músculo esquelético de SCD1 en el sujeto humano. El ejercicio crónico aeróbico influye en los niveles de proteína SCD1 en las personas. Un estudio que implica a atletas entrenados de resistencia y sujetos delgados sedentarios y obesos sedentarios fue conducido para evaluar diferencias del contenido en proteína SCD1.
La expresión de la proteína SCD1 era considerablemente más alto en atletas comparados con ambos grupos sedentarios. Pruebas también han manifestado que los atletas de resistencia han aumentado triglicéridos intramusculares (IMTG) y el ejercicio aumenta tanto la ß -oxidación como la lipogénesis dentro del músculo.
El papel de SCD1 en la Señalización de la Insulina y la Sensibilidad dentro del Músculo Esquelético
La SCD1de ratones knockout global (gKO) ha demostrado aumentos marcados en la señalización de la insulina en el tejido del hígado, adiposo, y el músculo. Los ratones gKO son ratones genéticamente son modificados para carecer de la expresión SCD1 en todos los tejidos. Rahman. et al. encontraron un aumentó en la activación glucógeno sintasa y la glucógeno fosforilasa, aumentó el contenido de GLUT4, y elevación en el receptor de insulina. Estos ratones también expusieron un aumento significativo de la activación (AMPK). Además, SCD1en ratones gKO muestran un aumento dramático de termogénesis, ß-oxidación, y la sensibilidad de insulina en el músculo esquelético y en otros tejidos.
SCD1 y Lipotoxicidad en el Músculo Esquelético
SFA puede causar stress celular e inflamación por varios mecanismos diferentes. SCD1 puede reducir la señalización proinflamatoria por aumento de la conversión SFA en MUFA.

Es generalmente aceptado que SFA, en particular el ácido palmítico y el ácido esteárico, es considerado lipotóxico en células y promueve la resistencia de insulina. El resultado de la lipotoxicidad puede provocar stress en el retículo endoplasmático (ER) y además inflamaci
ón, que es causada por la expresión citokinas. Los SFA de cadena larga activan el receptor (TLR4), que también causa la stress en el ER y una respuesta proinflamatoria. Sin embargo, la capacidad de ácido palmítico para inducir la resistencia de la insulina era notablemente inferior cuando TLR4 fue inhibido en rata.La alta expresión SCD1 puede ser beneficiosa con respecto a reducir SFA que puede activar TLR4. Mientras lo han mostrado que el aumentó de la expresión de SCD1 en células de músculo esquelético expuestas al ácido palmítico, lo rescata de la lipotoxicidad y la resistencia a la insulina por reducción de la inflamación y del stress del ER. Futuros estudios deberían ser conducidos para determinar si la expresión SCD1 puede reducir la activación TLR4 en el músculo esquelético.
Ácidos Grasos y Balance energético Influyen sobre la Expresión de SCD1en el Músculo Esquelético
A diferencia de la obesidad o el ejercicio, los ácidos grasos poliinsaturados (PUFA) disminuyen la expresión SCD1 en el músculo esquelético. Un estudio conducido por Sotavento et al. (37) divulgó que mRNA SCD1 estaba aumentado con una dieta alta en SFA y disminuido con dieta alta en PUFA. MUFA aparece disminuir la expresión SCD1 in vitro, aunque los resultados de cultivos celulares y en experimentos in vivo no sean concluyentes. Estas pruebas pueden indicar que la expresión SCD1 puede ser afectada por la presencia de MUFA y PUFAS y puede ser inhibida mediante feedback.
Papel del SCD1 en la Oxidación de Ácidos Grasos del Músculo Esquelético , Lipogénesis, y Esterificación
En atletas de resistencia se ha visto una alta expresión SCD1 y el contenido de IMTG, y elevación de la expresión de SCD1 puede promover un aumento de MUFA en el contenido de IMTG y la fluidez de membrana de la célula. No sea entendido completamente por qué aumentó los niveles de SCD1 en individuos obesos correlacionados positivamente con la resistencia a la insulina mientras que la alta expresión SCD1 en atletas es asociada con la sensibilidad de insulina. Esto parcialmente podría ser atribuido a una disminución de la ß-oxidación dentro del músculo esquelético como consecuencia de la obesidad, mientras que entrenamiento aeróbico en atletas demostró aumentó de la ß-oxidación.
El aumento de la ß-oxidación con la actividad física provocada en parte por un aumento de la activación del AMPK, que conduce a un aumento de la expresión de SCD1.

Los datos de ratones gKO SCD1 han mostrado aumentos de la activación de AMPK en fibras musculares oxidativas de tipo I , pero no fibras musculares de tipo II glucolíticas. Los investigadores especularon que el aumentó de la activación de AMPK en las fibras oxidativas puede inducir la ß-oxidación de ácidos grasos en ratones gKO SCD1. Un aumento de la carnitine palmitoyltransferase-1 (CPT-1) fue observado en las fibras oxidativas del tipo I en ratones gKO SCD1. Además, se ha visto que el consumo de O2 esta crónicamente elevado durante el ayuno y la alimentación en ratones gKO SCD1, que significa que el gasto de energía es más alto y pueden indicar una oxidación de ácidos grasos en estos animales. No sólo esta la ß-oxidación aumentada sino que la lipogénesis parece estar reducida también en ratones gKO SCD1.
En humanos, el ejercicio agudo aumentó SCD1 y aumentó la síntesis IMTG bajando DAG y reduciendo la inflamación. El aumento de la producción de MUFA parece aumentar la formación de TG y puede disminuir la lipotoxicidad y mejorar la señalización de la insulina.
La Expresión de SCD1 Regula la Síntesis de Ceramidas en el Músculo Esquelético
SCD1 puede tener un efecto fuerte sobre la composición ácidos grasos dentro del músculo esquelético. El aumento de la expresión SCD1 no sólo es beneficioso para la conversión de SFA en MUFA, también puede disminuir la acumulación ceramidas. De la síntesis novo de ceramidas en los humanos está regulado por seis ceramidas sintasas. En el músculo esquelético, la ceramida synthase-1, que sintetiza ceramidas con Stearoyl-CoA, es la forma predominante de ceramida sintasa. Otras sintasas sintetizan ceramidas con Palmitoyl-CoA que está también dentro del músculo esquelético. La alta actividad SCD1 reduce el flujo de SFA hacia la ceramida sintasa, convirtiéndolos (SFA) en MUFA.
A la inversa, disminuyendo la actividad SCD1 permite la acumulación de MUFA, causando un aumento de las vías de síntesis de ceramidas y la acumulación ceramidas en el tejido del músculo.
Table 2.
Los efectos de la composición de ácidos grasos en la expresión de SCD1 en el músculo esquelético
| Organism/Model | SCD1 Expression | Skeletal Muscle Fatty Acid Composition | Pertinent Findings from Study |
|---|---|---|---|
| Obese and lean human females (29) | Higher in obese than in lean subjects | Obese had higher oleic acid, lower palmitic acid, and lower stearic acid than lean subjects | Fivefold higher fasting insulin levels in obese than in lean subjects |
| Healthy males with impaired glucose tolerance on rosiglitazone therapy for eight weeks (41) | Increased posttherapy | Reduction in stearic acid and increases in palmitoleate acid, oleic acid, and vaccenic acid | Decreased glucose and insulin during oral glucose tolerance testing |
| Healthy human males consuming a high-fructose diet for four weeks (35) | Increased postdiet | No change in intramyocellular lipids | Decrease in skeletal muscle GLUT4 and increase in fasting plasma glucose, leptin, TG, and VLDL-TG, but no change in insulin-mediated glucose disposal |
| Healthy premenopausal humans engaging in one bout of exercise (70) | Increased after one bout of exercise | Increase in TG, decrease in DAG, and decrease in ceramides | Decrease in inflammatory pathways and increased insulin sensitivity and fatty acid oxidation |
| Male athletes and sedentary men (4) | Higher in athletes | Higher total TG but lower saturated DAG in athletes | Twofold higher insulin sensitivity in athletes |
| Human athletes, sedentary normal-weight humans, and sedentary obese humans (3) | SCD1 highest in athletes | Ceramides were highest in the sedentary obese group, while DAG was highest in athletes, and TG were highest in both athletes and sedentary obese | Peripheral insulin sensitivity highest in athletes, followed by lean sedentary, then sedentary obese, and mitochondrial volume density was highest in athletes |
| Male Zucker diabetic fatty rats and male lean Zucker rats (84) | Higher in Zucker diabetic fatty rats than in lean Zucker rats | Zucker diabetic fatty rats had higher palmitoleoyl-CoA than lean Zucker rats | Zucker diabetic fatty rats displayed hyperinsulinemia, hyperglycemia, hypertriglyceridemia, hyperleptinemia, and elevated Hb A1c levels. |
| Male Sprague-Dawley rats fed an eight-week high-SFA diet, high-PUFA diet, or control diet for eight weeks (37) | Highest in high-SFA diet | TG (total and percent unsaturated fatty acids) were highest in high-PUFA diet, while total DAG and percent saturated fatty acids within TG were highest in high-SFA diet | Compared to control diet, high-SFA diet decreased glucose tolerance, while high-PUFA diet increased insulin sensitivity |
| Male Wistar rats performing a six-week treadmill program compared to sedentary controls (18) | Increased in soleus in exercise-trained rats but remained unchanged in EDL | FFA, DAG, and TG increased in soleus of exercise-trained rats but remained unchanged in EDL | PPARγ, PPARδ, acetyl-CoA carboxylase, fatty acid synthase, GPAT, and DGAT all increased in the soleus of exercise trained rats |
- DAG, diacylglycerol; TG, triglyceride; SFA, saturated fatty acid; PPAR, peroxisome proliferator-activated receptor; GPAT, glycerol-3-phosphate acyltransferase; DGAT, acyl-CoA:diaclyglycerol acyltransferase.
Se ha demostrado que la síntesis de ceramidas en el músculo esquelético humano por la exposición a ácido palmítico puede provocar resistencia a la insulina. Un estudio que implica a atletas delgados, sujetos sedentarios delgados y participantes obesos donde se analizó la composición de ácidos grasos, se descubrió que el total, de ceramidas saturadas e insaturadas que fueron encontradas dentro del músculo esquelético, fueron considerablemente más altas en el grupo obesos que en el grupo de delgados y atletas. Además, los niveles de SCD1 en el estudio mostraron una relación positiva con la sensibilidad de la insulina, mientras que fueron observadas correlaciones negativas con el total, de ceramidas saturadas e insaturadas. Un descubrimiento sorprendente del estudio consistió en que el DAG, que, como se piensa, promueve la resistencia de la insulina del músculo esquelético, era más alto en los atletas que en el grupo de sedentarios delgados y obesos, los investigadores principales del estudio para asumir que la paradoja " del atleta " puede ser aún más complicada que al principio se esperada (p. ej., no exclusivamente restringida a TG)
El contenido total de DAG no puede ser perjudicial en sí mismo, pero en cambio ciertas especies de DAG pueden ser lipotóxicas. En un estudio in vitro que se usan miocitos de ratón expuesto a SFA, demostraron que la acumulación ceramidas, y no la acumulación DAG , perjudicada las vías señalizadoras de la insulina.

















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}