PROGRAMA DE NUTRICIÓN COMUNITARIA PARA ENFERMOS DE CANCER
<iframe src="https://docs.google.com/presentation/d/1AHAlXs9qMCihznSopZFfL9WeFn81cveOBm_rjzLT7vo/embed?start=false&loop=false&delayms=3000" frameborder="0" width="960" height="749" allowfullscreen="true" mozallowfullscreen="true" webkitallowfullscreen="true"></iframe>
lunes, 25 de febrero de 2013
domingo, 17 de febrero de 2013
Cambios en la Prueba de sobrecarga de glucosa en obesos tras entrenamiento físico
Los resultados del trabajo de (Bruce. Endurance training in obese humans improves glucose tolerance and mitochondrial fatty acid oxidation and alters muscle lipid content.
Am J Physiol Endocrinol Metab 291: E99–E107, 2006) se puede observar claramente como la glucosa ( P < 0.05) y la insulina ( P = 0.01) disminuyen sus valores posejercicio a una prueba de sobrecarga de glucosa. El ejercicio realizado fue 5 veces x semana durante 8 semanas - 60 min al 60-70% del VO2 máximo. Incremento de la actividad de la CPT 1 en un 250% ( P 0.001) con el entrenamiento y se hizo menos sensible a la inhibición por Malonyl-CoA. Esto fue asociado con un aumento de la oxidación de Acidos Grasos mitocondrial (120%, P < 0.001)

Bruce. Endurance training in obese humans improves glucose tolerance and mitochondrial fatty acid oxidation and alters muscle lipid content.
Am J Physiol Endocrinol Metab 291: E99–E107, 2006
Am J Physiol Endocrinol Metab 291: E99–E107, 2006) se puede observar claramente como la glucosa ( P < 0.05) y la insulina ( P = 0.01) disminuyen sus valores posejercicio a una prueba de sobrecarga de glucosa. El ejercicio realizado fue 5 veces x semana durante 8 semanas - 60 min al 60-70% del VO2 máximo. Incremento de la actividad de la CPT 1 en un 250% ( P 0.001) con el entrenamiento y se hizo menos sensible a la inhibición por Malonyl-CoA. Esto fue asociado con un aumento de la oxidación de Acidos Grasos mitocondrial (120%, P < 0.001)
Se han propuesto que las intervenciones que conducen a una capacidad aumentada de oxidación de Acidos Grasos para el músculo esquelético, comoel entrenamiento de resistencia, puedan mejorar la acción de insulina por una reducción de lípidos intramusculares. Sin embargo, muy poco se sabe en cuanto al efecto del entrenamiento resistencia sobre músculo esquelético.
Estos cambios de lípidos intramusculares fueron correlacionados con la mejora observada de la tolerancia de glucosa. La CPT1 está considerada el paso limitante que interviene la oxidación de Acidos Grasos Saturados. La CPT1 está reducida la actividad en el músculo esquelético de individuos obesos y probablemente contribuye a las tasas disminuidas de oxidación de Acidos Grasos en la obesidad.
Bruce. Endurance training in obese humans improves glucose tolerance and mitochondrial fatty acid oxidation and alters muscle lipid content.
Am J Physiol Endocrinol Metab 291: E99–E107, 2006
Adaptación del Músculo Esquelético en Niños Adolescentes al entrenamiento aerobico y de velocidad
El estudio fue investigar los efectos de 3 meses de entrenamiento aeróbico y de velocidad sobre el área de la fibras y de la actividad glucolítica (PFK fosfofructokinasa) y oxidativa (SDH succinato deshidrogenasa) del vasto lateral en niños adolescentes. La actividad enzimática fue también determinada luego de 6 meses de desentrenamiento.
El entrenamiento aeróbico generó un aumento significativo en el VO2 max. (58.4 – 64.3 mlxkgxmin), un aumento del área de la fibra ST y FTa (6.0 – 7.3 y 8.0 – 10.4 µm2 x 103) respectivamente y en la actividad de la SDH (6.4 – 9.1 IU).
Luego del desentrenamiento el VO2 max y la actividad de la SDH retornaron a los niveles de preentrenamiento. El entrenamiento de velocidad resultó en un aumento significativo solo en la actividad de la PFK (28.1 – 33.9 IU) el cual también disminuyó luego del periodo
Doce sujetos adolescentes masculinos de edades entre 16-17 años, fueron asignados al azar a dos grupos de entrenamiento bajo supervisión de 4 veces por semana durante 3 meses. Se obtuvieron consentimientos informados de cada sujeto firmados por sus padres. El grupo de entrenamiento aeróbico (END) realizó trote, el cual fue progresivamente incrementado desde 2 pasadas de 10 minutos a 2 pasadas de 30 minutos con 5 minutos de pausa entre pasadas. Esto se realizó a velocidades que requerían del 60 al 70% de la máxima frecuencia cardiaca al comienzo del entrenamiento alcanzando del 80-90% al finalizar el entrenamiento. El grupo de entrenamiento de la velocidad (SP) realizó carreras intervaladas de 50 a 250 metros y ocasionalmente carreras en escalera cuesta arriba. El tiempo de entrenamiento utilizado en cada grupo fue el mismo. Las siguientes evaluaciones fueron realizadas antes y después de 3 meses de entrenamiento; algunas de ellas fueron repetidas luego de 6 meses de desentrenamiento.
Resultados
El entrenamiento resultó en un incremento significativo en VO2 max (lxmin) para END (12%) y SP (10%) (tabla 1).

Tabla 1. Datos pre, post y desentrenamiento
Valores en media y desvío estándar
* = diferencia significativa entre pre y post entrenamiento (p<0.05).
† = diferencia significativa entre post y desentrenamiento (p<0.05) usando ANOVA mediciones repetidas.
SP= grupo entrenamiento velocidad; END= grupo entrenamiento aeróbico; N=6 sujetos por grupo
El descenso en la máxima frecuencia cardiaca fue significante en el grupo END. Durante el ejercicio submáximo (0% inclinación – 7 mph) la frecuencia cardiaca fue (p<0.05) significativamente más baja tanto para el grupo SP (168 ± 12 a 163 ± 11 latxmin) como para el grupo END (171 ± 12 a 158 ± 12 latxmin). Luego de 6 meses de desentrenamiento el VO2 max no fue diferente significativamente de los valores de antes del entrenamiento pero si fueron diferentes de los valores de luego del periodo de entrenamiento.
El grupo END mostró aumentos significativos en el área de las fibras musculares ST, FTa y FTc, mientras que no fue significativo para el grupo que entrenó velocidad (tabla 2)

Tabla 2. Efectos del entrenamiento sobre el área y la distribución de la fibra muscular esquelética.
Para descripción de terminos ver tabla 1 y texto.
* = diferencia significativa entre pre y post entrenamiento (p<0.05).
La distribución de las fibras permaneció sin cambios en ambos grupos. El grupo END mostró un 42% de aumento en la actividad de la SDH en el vasto lateral (tabla 3 – figura 1) y ningún cambio en la actividad de la PFK (tabla 3 – figura 2)
Por otro lado, el grupo SP mostró un aumento del 21% en la actividad de la PFK sin ninguna modificación en la SDH. Luego de 6 meses de desentrenamiento, la actividad de la SDH en el vasto lateral en el grupo END y la actividad de la PFK en el grupo SP retornaron a valores más bajos que antes del entrenamiento aunque no fue significativo. La actividad de la SDH en el grupo SP fue significativamente más baja (p<0.05) luego del desentrenamiento comparado con los valores antes y después del entrenamiento.

Tabla 3. Efectos de entrenamiento y desentrenamiento sobre la actividad de las enzimas musculares en adolescentes.
Para descripción de terminos ver tabla 1 y texto.
IU= µmoles x gr x min
* = diferencia significativa entre pre y post entrenamiento (p<0.05).
† = diferencia significativa entre post y desentrenamiento (p<0.05) usando ANOVA mediciones repetidas.
SP= grupo entrenamiento velocidad; END= grupo entrenamiento aeróbico; N=6 sujetos por grupo


El entrenamiento aeróbico generó un aumento significativo en el VO2 max. (58.4 – 64.3 mlxkgxmin), un aumento del área de la fibra ST y FTa (6.0 – 7.3 y 8.0 – 10.4 µm2 x 103) respectivamente y en la actividad de la SDH (6.4 – 9.1 IU).
Luego del desentrenamiento el VO2 max y la actividad de la SDH retornaron a los niveles de preentrenamiento. El entrenamiento de velocidad resultó en un aumento significativo solo en la actividad de la PFK (28.1 – 33.9 IU) el cual también disminuyó luego del periodo
Doce sujetos adolescentes masculinos de edades entre 16-17 años, fueron asignados al azar a dos grupos de entrenamiento bajo supervisión de 4 veces por semana durante 3 meses. Se obtuvieron consentimientos informados de cada sujeto firmados por sus padres. El grupo de entrenamiento aeróbico (END) realizó trote, el cual fue progresivamente incrementado desde 2 pasadas de 10 minutos a 2 pasadas de 30 minutos con 5 minutos de pausa entre pasadas. Esto se realizó a velocidades que requerían del 60 al 70% de la máxima frecuencia cardiaca al comienzo del entrenamiento alcanzando del 80-90% al finalizar el entrenamiento. El grupo de entrenamiento de la velocidad (SP) realizó carreras intervaladas de 50 a 250 metros y ocasionalmente carreras en escalera cuesta arriba. El tiempo de entrenamiento utilizado en cada grupo fue el mismo. Las siguientes evaluaciones fueron realizadas antes y después de 3 meses de entrenamiento; algunas de ellas fueron repetidas luego de 6 meses de desentrenamiento.
Resultados
El entrenamiento resultó en un incremento significativo en VO2 max (lxmin) para END (12%) y SP (10%) (tabla 1).
Tabla 1. Datos pre, post y desentrenamiento
Valores en media y desvío estándar
* = diferencia significativa entre pre y post entrenamiento (p<0.05).
† = diferencia significativa entre post y desentrenamiento (p<0.05) usando ANOVA mediciones repetidas.
SP= grupo entrenamiento velocidad; END= grupo entrenamiento aeróbico; N=6 sujetos por grupo
El descenso en la máxima frecuencia cardiaca fue significante en el grupo END. Durante el ejercicio submáximo (0% inclinación – 7 mph) la frecuencia cardiaca fue (p<0.05) significativamente más baja tanto para el grupo SP (168 ± 12 a 163 ± 11 latxmin) como para el grupo END (171 ± 12 a 158 ± 12 latxmin). Luego de 6 meses de desentrenamiento el VO2 max no fue diferente significativamente de los valores de antes del entrenamiento pero si fueron diferentes de los valores de luego del periodo de entrenamiento.
El grupo END mostró aumentos significativos en el área de las fibras musculares ST, FTa y FTc, mientras que no fue significativo para el grupo que entrenó velocidad (tabla 2)
Tabla 2. Efectos del entrenamiento sobre el área y la distribución de la fibra muscular esquelética.
Para descripción de terminos ver tabla 1 y texto.
* = diferencia significativa entre pre y post entrenamiento (p<0.05).
La distribución de las fibras permaneció sin cambios en ambos grupos. El grupo END mostró un 42% de aumento en la actividad de la SDH en el vasto lateral (tabla 3 – figura 1) y ningún cambio en la actividad de la PFK (tabla 3 – figura 2)
Por otro lado, el grupo SP mostró un aumento del 21% en la actividad de la PFK sin ninguna modificación en la SDH. Luego de 6 meses de desentrenamiento, la actividad de la SDH en el vasto lateral en el grupo END y la actividad de la PFK en el grupo SP retornaron a valores más bajos que antes del entrenamiento aunque no fue significativo. La actividad de la SDH en el grupo SP fue significativamente más baja (p<0.05) luego del desentrenamiento comparado con los valores antes y después del entrenamiento.
Tabla 3. Efectos de entrenamiento y desentrenamiento sobre la actividad de las enzimas musculares en adolescentes.
IU= µmoles x gr x min
* = diferencia significativa entre pre y post entrenamiento (p<0.05).
† = diferencia significativa entre post y desentrenamiento (p<0.05) usando ANOVA mediciones repetidas.
SP= grupo entrenamiento velocidad; END= grupo entrenamiento aeróbico; N=6 sujetos por grupo
Discusión
Los presentes resultados confirman los hallazgos encontrados en adolescentes y posiblemente en niños post púberes. Ambos grupos de entrenamiento han mostrado incrementos en el VO2 max (2-18). Los resultados de este estudio indican una respuesta enzimática es específica para cada tipo de entrenamiento a pesar de un aumento similar en el VO2 max. El grupo END o de entrenamiento continuo aumentó la actividad enzimática y el grupo SP o de potencia incrementó la actividad glucolítica. Los cambios en el VO2 max y la actividad enzimática oxidativa puede ocurrir separadamente como ha sido demostrado por Henriksson y Reitman (15). Una adaptación específica fue mostrada por el incremento con entrenamiento aeróbico en el área de las fibras ST y FTa. Esto sugirió que el entrenamiento aeróbico estimula unidades motoras particulares que generó hipertrofia (3-10-14). Sin embargo el grado de tensión generado durante el entrenamiento de velocidad fue adecuado para estimular la vía glucolítica anaeróbica aunque el tiempo y la tensión generada no fue suficiente para aumentar el tamaño de la célula. La adaptación de la actividad enzimática oxidativa en el musculo esquelético en adolescentes es similar a la observada en adultos (10-12-13-17-19). Sin embargo los bajos valores de PFK y la falla de hipertrofiar las fibras FTb con el entrenamiento de velocidad difieren de los hallazgos encontrados en adultos. Finalmente el crecimiento parece no influenciar los presentes resultados por que el peso y la talla no se modificaron durante los 3 meses de entrenamiento y las enzimas no cambiaron significativamente luego de 9 meses de haber comenzado el estudio con la excepción de una actividad disminuida en el grupo de entrenamiento de velocidad.
Referencias
1. Allen TH, MT Peng, KP Cheng, TF Hyang, C Chang y HS Fang (1956). Prediction of total adiposity from skinfolds and curvilinear relationship between external and internal adiposity. Metabolism. 5:346-352
2. Astrand PO (1952). Experimental studies of physical working capacity in relation to sex and age. Copenhagen: Munksgaard. P 56
3. Bell RD, JD MacDougall, R Billeter and H Howald (1980). Muscle fiber types and morphometric analysis of skeletal muscle in six year old children. Med. Sci Sports Exercise. 12:28-31
4. Bergmeyer U (1974). Methods of enzymatic analysis. New York. Academic Press, p 451
5. Bergstrom J (1962). Muscle electrolytes in man. Scand J. Clin. Lab. Invest. Supplement. 68:1-110
6. Dubowitz U and MH Brooke (1973). Muscle biopsy: a modern approach. London WB Sounders, p 475
7. Eriksson BO (1972). Physical training, oxygen supply and muscle metabolism in 11-13 year old boys. Acta Physiol. Scand. Supplement. 384:1-48
8. Eriksson BO, PD Gollnick and B Saltin (1973). Muscle metabolism and enzyme activities after training in boys 11-13 years old. Acta Physiol. Scand. 87:485-497
9. Eriksson BO, J Karlsson and B Saltin (1971). Muscle metabolism during exercise in 13 years old boys. Acta Paediatr. Scand. 60 Supplement. 217, 57-63
10. Essen B, E Jansson, J Henriksson, AW Taylor and B Saltin (1975). Metabolic characteristics of fibre type in human skeletal muscle. Acta Physiol. Scand. 95:153-165
11. Ferguson RJ, J Charlebois, AW Taylor et al (1979). Peripheral adaptations with training in patients with angina pectoris. Circulation 60 Supplement II. Abstract, 235
12. Gollnick PD, RB Armstrong, B Saltin, CW Saubert, WL Sembrowich and RE Shepherd (1973). Effect of training on enzyme activity and fibre composition of human skeletal muscle. J. Appl. Physiol. 34:107-111
13. Gollnick PD, RB Armstrong, CW Saubert, K Piehl and B Saltin (1972). Enzyme activity and fibre composition in skeletal muscle of untrained and trained men. J. Appl. Physiol. 33:312-319
14. Henriksson J and JS Reitman (1976). Quantitative measures of enzyme activities in type I and type II fibers of man after training. Acta Physiol. Scand. 97:392-397
15. Henriksson J and JS Reitman (1977). Time course of changes in human skeletal muscle succinate dehydogenase and cytochrome oxidase activities and maximal oxygen uptake with physical activity and inactivity. Acta Physiol. Scand. 99:91-97
16. Jansson E and L Kaisjer (1977). Muscle adaptation to extreme endurance training in man. Acta Physiol. Scand. 100:315-324
17. Karlsson J (1971). Lactate and phosphagen concentrations in working muscle man. Acta Physiol. Scand. Supplement. 358:1-72
18. Knuttgen HG, LO Norder, B Ollander and B Saltin (1973). Physiological conditioning through internal training with young male adults. Med. Sci. Sports. 5:220-226
19. Komi PV and J Karlsson (1978). Skeletal muscle fibre types enzyme activities and physical performance in young males and females. Acta Physiol. Scand. 103:210-218
20. Leger L, V Seliger and L Brassard (1980). Backward extrapolation of VO2 max. values from the O2 recovery curve. Med. Sci. Sports Exercise. 12:24-27
21. Novikoff AB, WY Shin and J Drucker (1961). Mitochondrial localization of oxidative enzymes staining results with two tatrazolium salts. J. Biophys. Biochem. Cytol. 9:47-61
22. Saltin B (1973). Metabolic fundamentals in exercise. Med. Sci. Sports. 5:137-146
23. Saltin B, K Nazar, DL Costill et al (1976). The nature of the training response: peripheral and central adaptations to one legged exercise. Acta Physiol. Scand. 96:289-305
24. Sembrowich WL, MB Knudson and PD Gollnick (1977). Muscle metabolism and cardiac function in the myopathic hamster following training. J. Appl. Physiol. 43:936-941
25. Taylor AW, S Lavoie, G Lemieux, C Dufresne, JS Skinner and J Vallee (1978). Effects of endurance training on the area and enzyme activities of skeletal muscle of French Canadians. In: 3rd International Symposium on biochemistry of exercise. F Landry and WAR Orban (Eds). Miami: Sumposia Specialist, pp 267-278
26. Thorstensson A, B Sjodin and J Karlsson (1975). Enzyme activities and muscle after sprint training in man. Acta Physiol. Scand. 94:313-318
2. Astrand PO (1952). Experimental studies of physical working capacity in relation to sex and age. Copenhagen: Munksgaard. P 56
3. Bell RD, JD MacDougall, R Billeter and H Howald (1980). Muscle fiber types and morphometric analysis of skeletal muscle in six year old children. Med. Sci Sports Exercise. 12:28-31
4. Bergmeyer U (1974). Methods of enzymatic analysis. New York. Academic Press, p 451
5. Bergstrom J (1962). Muscle electrolytes in man. Scand J. Clin. Lab. Invest. Supplement. 68:1-110
6. Dubowitz U and MH Brooke (1973). Muscle biopsy: a modern approach. London WB Sounders, p 475
7. Eriksson BO (1972). Physical training, oxygen supply and muscle metabolism in 11-13 year old boys. Acta Physiol. Scand. Supplement. 384:1-48
8. Eriksson BO, PD Gollnick and B Saltin (1973). Muscle metabolism and enzyme activities after training in boys 11-13 years old. Acta Physiol. Scand. 87:485-497
9. Eriksson BO, J Karlsson and B Saltin (1971). Muscle metabolism during exercise in 13 years old boys. Acta Paediatr. Scand. 60 Supplement. 217, 57-63
10. Essen B, E Jansson, J Henriksson, AW Taylor and B Saltin (1975). Metabolic characteristics of fibre type in human skeletal muscle. Acta Physiol. Scand. 95:153-165
11. Ferguson RJ, J Charlebois, AW Taylor et al (1979). Peripheral adaptations with training in patients with angina pectoris. Circulation 60 Supplement II. Abstract, 235
12. Gollnick PD, RB Armstrong, B Saltin, CW Saubert, WL Sembrowich and RE Shepherd (1973). Effect of training on enzyme activity and fibre composition of human skeletal muscle. J. Appl. Physiol. 34:107-111
13. Gollnick PD, RB Armstrong, CW Saubert, K Piehl and B Saltin (1972). Enzyme activity and fibre composition in skeletal muscle of untrained and trained men. J. Appl. Physiol. 33:312-319
14. Henriksson J and JS Reitman (1976). Quantitative measures of enzyme activities in type I and type II fibers of man after training. Acta Physiol. Scand. 97:392-397
15. Henriksson J and JS Reitman (1977). Time course of changes in human skeletal muscle succinate dehydogenase and cytochrome oxidase activities and maximal oxygen uptake with physical activity and inactivity. Acta Physiol. Scand. 99:91-97
16. Jansson E and L Kaisjer (1977). Muscle adaptation to extreme endurance training in man. Acta Physiol. Scand. 100:315-324
17. Karlsson J (1971). Lactate and phosphagen concentrations in working muscle man. Acta Physiol. Scand. Supplement. 358:1-72
18. Knuttgen HG, LO Norder, B Ollander and B Saltin (1973). Physiological conditioning through internal training with young male adults. Med. Sci. Sports. 5:220-226
19. Komi PV and J Karlsson (1978). Skeletal muscle fibre types enzyme activities and physical performance in young males and females. Acta Physiol. Scand. 103:210-218
20. Leger L, V Seliger and L Brassard (1980). Backward extrapolation of VO2 max. values from the O2 recovery curve. Med. Sci. Sports Exercise. 12:24-27
21. Novikoff AB, WY Shin and J Drucker (1961). Mitochondrial localization of oxidative enzymes staining results with two tatrazolium salts. J. Biophys. Biochem. Cytol. 9:47-61
22. Saltin B (1973). Metabolic fundamentals in exercise. Med. Sci. Sports. 5:137-146
23. Saltin B, K Nazar, DL Costill et al (1976). The nature of the training response: peripheral and central adaptations to one legged exercise. Acta Physiol. Scand. 96:289-305
24. Sembrowich WL, MB Knudson and PD Gollnick (1977). Muscle metabolism and cardiac function in the myopathic hamster following training. J. Appl. Physiol. 43:936-941
25. Taylor AW, S Lavoie, G Lemieux, C Dufresne, JS Skinner and J Vallee (1978). Effects of endurance training on the area and enzyme activities of skeletal muscle of French Canadians. In: 3rd International Symposium on biochemistry of exercise. F Landry and WAR Orban (Eds). Miami: Sumposia Specialist, pp 267-278
26. Thorstensson A, B Sjodin and J Karlsson (1975). Enzyme activities and muscle after sprint training in man. Acta Physiol. Scand. 94:313-318
Medicine and Science in Sports and Exercise. Vol 14, N°6, pp 453-456, 1982. Mario Fournier, Joe Ricci, Albert Taylor, Ronald Feeguson, Richard Montpetit y Bernard Chaitman.
lunes, 11 de febrero de 2013
NUTRIGENÉTICA: Los genes IKKE y TBK1 jugan un papel crucial para mantener el equilibrio metabólico
Un antiguo fármaco para el asma puede señalar el camino a nuevos tratamientos para la diabetes y la obesidad.
 "Una de las razones por la que las dietas son tan efectivas a la hora de perder peso para algunas personas es que sus cuerpos se ajustan a las calorías también mediante la reducción de su metabolismo, por lo que están defendiendo su peso corporal. Amlexanox parece modificar la respuesta metabólica al almacenamiento excesivo de calorías en ratones", explica Alan Saltiel, director del Instituto de Ciencias de la Vida.
"Una de las razones por la que las dietas son tan efectivas a la hora de perder peso para algunas personas es que sus cuerpos se ajustan a las calorías también mediante la reducción de su metabolismo, por lo que están defendiendo su peso corporal. Amlexanox parece modificar la respuesta metabólica al almacenamiento excesivo de calorías en ratones", explica Alan Saltiel, director del Instituto de Ciencias de la Vida.

http://www.europapress.es/salud/noticia-antiguo-farmaco-asma-puede-senalar-camino-nuevos-tratamientos-diabetes-obesidad-20130210190807.html
Investigadores del Instituto de Ciencias de la Vida de la Universidad de Michigan (Estados Unidos) han descubierto que amlexanox, un fármaco sin patente que actualmente se receta para el tratamiento del asma y otras aplicaciones, también revierte la obesidad, la diabetes y el hígado graso en ratones, según publica en su edición de este domingo la revista 'Nature Medicine'.
 "Una de las razones por la que las dietas son tan efectivas a la hora de perder peso para algunas personas es que sus cuerpos se ajustan a las calorías también mediante la reducción de su metabolismo, por lo que están defendiendo su peso corporal. Amlexanox parece modificar la respuesta metabólica al almacenamiento excesivo de calorías en ratones", explica Alan Saltiel, director del Instituto de Ciencias de la Vida.
"Una de las razones por la que las dietas son tan efectivas a la hora de perder peso para algunas personas es que sus cuerpos se ajustan a las calorías también mediante la reducción de su metabolismo, por lo que están defendiendo su peso corporal. Amlexanox parece modificar la respuesta metabólica al almacenamiento excesivo de calorías en ratones", explica Alan Saltiel, director del Instituto de Ciencias de la Vida.
Diferentes formulaciones de amlexanox están actualmente prescritas para tratar el asma en Japón y las aftas bucales en Estados Unidos. Saltiel y sus colegas de la Universidad de Michigan probaron si amlexanox es útil para el tratamiento de la obesidad y la diabetes en los seres humanos y se puede desarrollar un nuevo compuesto centrado en esta droga que optimice su fórmula.
El estudio confirma y amplía la noción de que los genes IKKE y TBK1 jugan un papel crucial para mantener el equilibrio metabólico, un descubrimiento publicado por el laboratorio Saltiel en 2009 en la revista 'Cell'. "Amlexanox parece que funciona en ratones, inhibiendo IKKE y TBK1 que creemos que juntos actúan como una especie de freno en el metabolismo. Al soltar el freno, amlexanox parece liberar el sistema metabólico para quemar más y posiblemente almacenar menos la energía", dijo Saltiel.
Con el uso de alto rendimiento químico en el Centro de LSI de Genómica Química para buscar compuestos que inhiben IKKE y TBK1, los investigadores probaron el medicamento fuera de patente amlexanox y demostraron que tiene efectos beneficiosos en ratones obesos tanto genética como dietéticamente inducidos. El producto químico redujo el peso de los ratones obesos y revirtió los problemas metabólicos relacionados como la diabetes y el hígado graso.
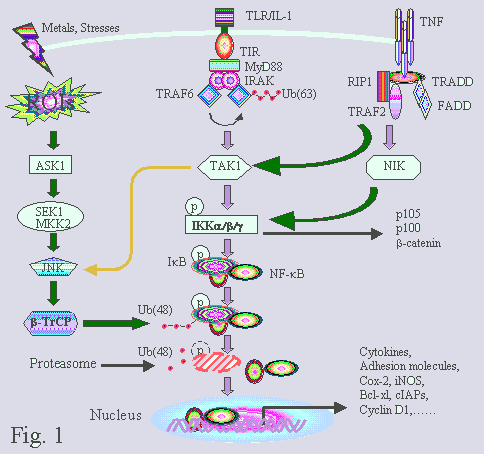
"Estos estudios nos dicen que, al menos en ratones, la vía IKKE/TBK1 juega un papel importante en la defensa de peso corporal mediante el aumento y la disminución de almacenamiento y quema de calorías y que, mediante la inhibición de que la vía con un compuesto, puede aumentar el metabolismo e inducir la pérdida de peso, revertir la diabetes y disminuir el hígado graso", resumió Saltiel.
Sin embargo, los investigadores no saben todavía si los seres humanos responden con la misma vía o si el descubrimiento de la eficacia amlexanox en ratones puede conducir a un compuesto que es seguro y eficaz para el tratamiento de la obesidad y la diabetes en los seres humanos. "Vamos a trabajar duro en esto", concluye el investigador principal.
http://www.europapress.es/salud/noticia-antiguo-farmaco-asma-puede-senalar-camino-nuevos-tratamientos-diabetes-obesidad-20130210190807.html
jueves, 7 de febrero de 2013
NUTRIGENÉTICA:La hipometilación del gen que codifica IFG2 (factor de crecimiento insulínico tipo 2)
La obesidad paterna eleva el riesgo de cáncer del niño
La dieta materna y el peso pueden influir en la salud del niño antes del nacimiento. Sin embargo, según un trabajo que se publica hoy en BMC Medicine, la obesidad paterna también tiene impacto.
La hipometilación del gen que codifica IFG2 (factor de crecimiento insulínico tipo 2) en los recién nacidos se correlaciona con un riesgo aumentado de desarrollar cáncer a lo largo de la vida. Así, en los niños con padres varones obesos, se produce una reducción en la cantidad de ADN de la metilación de IGF2 en las células fetales aisladas en el cordón umbilical.
Los investigadores, del Duke University Hospital, en el Reino Unido, recogieron información sobre el peso de los padres y la compararon con los datos epigenéticos de los niños. "Las marcas epigenéticas se reprograman cuando el esperma y los óvulos se forman. No obstante, parece que la nutrición, el estilo de vida y el entorno de los padres tienen un efecto directo en la salud y desarrollo del niño", explica Catherine Hoyo, una de las autoras.
Diariomedico.com
La hipometilación del gen que codifica IFG2 (factor de crecimiento insulínico tipo 2) en los recién nacidos se correlaciona con un riesgo aumentado de desarrollar cáncer a lo largo de la vida. Así, en los niños con padres varones obesos, se produce una reducción en la cantidad de ADN de la metilación de IGF2 en las células fetales aisladas en el cordón umbilical.
Los investigadores, del Duke University Hospital, en el Reino Unido, recogieron información sobre el peso de los padres y la compararon con los datos epigenéticos de los niños. "Las marcas epigenéticas se reprograman cuando el esperma y los óvulos se forman. No obstante, parece que la nutrición, el estilo de vida y el entorno de los padres tienen un efecto directo en la salud y desarrollo del niño", explica Catherine Hoyo, una de las autoras.
Diariomedico.com
miércoles, 6 de febrero de 2013
PREVENCIÓN DE ENFERMEDADES NEURODEGENERATIVAS
<iframe src="https://docs.google.com/file/d/0B0LYxjcI1XGpTnhSOUxYTTBXUzQ/preview" width="640" height="480"></iframe>
<iframe src="https://docs.google.com/file/d/0B0LYxjcI1XGpTnhSOUxYTTBXUzQ/preview" width="640" height="480"></iframe>
Constante de Tiempo (τ) del Consumo de Oxígeno
Constante de Tiempo (τ) del Consumo de Oxígeno
Con la introducción de un estímulo de ejercicio constante, el consumo de oxígeno se incrementa con un patrón predecible que describe convenientemente la cinética de primer orden con una función exponencial (Ver Figura 1).
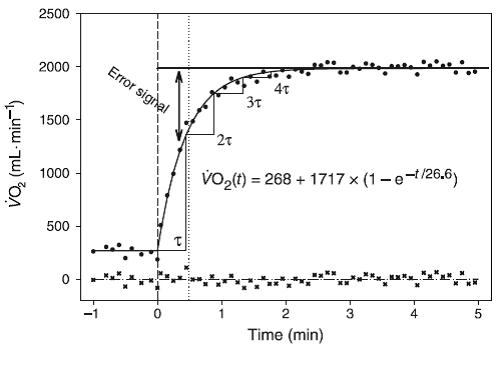
Figura 1.Conjunto hipotético de datos generados para mostrar algunos puntos clave relacionados con el ajuste de los datos de la cinética del VO2. En los primeros 30 s (entre la línea vertical a rayas y la línea punteada), la constante tau (τ) = 24 s, pero para todos los tiempos luego de esto, τ = 36 s.
Existe cierto debate acerca de la naturaleza exacta de esta función y si existe o no retrasos en el tiempo o componentes exponenciales múltiples. Sin embargo, para fines prácticos (i.e., pruebas de ejercicio para la valoración del rendimiento o la valoración clínica), el incremento en la cinética del consumo de oxígeno con el comienzo del ejercicio puede ser convenientemente descrito por una función mono-exponencial con una constante de tiempo (τVO2). Esta constante de tiempo se define en la ecuación estándar de una función exponencial.
VO2(t) = δVO2 (1 – e(t/τ))
Donde VO2(t) es el consumo de oxígeno a un tiempo (t) dado; δVO2 es el incremento total en el consumo de oxígeno y e es la base natural de los logaritmos. En base al modelo monoexponencial, luego de una constante de tiempo (τVO2), el incremento del VO2 será de ~ 63% del incremento total hasta un nuevo valor en estado estable, y luego de cuatro constantes de tiempo, el incremento en el VO2 será del 98% del incremento total.
La constante de tiempo τVO2 es importante debido a que, conjuntamente con el VO2máx, η y el umbral ventilatorio, es uno de los parámetros fundamentales de la función aeróbica. Por encima del umbral ventilatorio, la determinación de τVO2 es más compleja debido al continuo incremento en el VO2; incluso durante un ejercicio con carga constante.
La mejor forma de derivar la constante τVO2es a partir de un test de ejercicio a carga constante, en donde la entrada en calor es seguida por la imposición de una carga de trabajo de intensidad moderada por al menos 6 minutos. Durante este tipo de test, se espera que el VO2se incremente hasta un nuevo estado estable en aproximadamente 3 minutos. Se curva de ajuste exponencial se utiliza para derivar τVO2. Alternativamente, si se deriva el déficit de oxígeno (VO2def) utilizando métodos matemáticos más complejos, entonces τVO2 puede calcularse a partir de la siguiente ecuación:
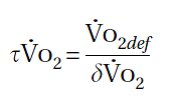
Siendo τVO2 una constante de tiempo, sus unidades son segundos.
El valor normal de τVO2 derivado de acuerdo con la ecuación presentada, en jóvenes sedentarios saludables es de 38 s con una desviación estándar de 5 s. La τVO2 se reduce con el entrenamiento y se ha observado que puede ser tan baja como de 20 s en atletas. La τVO2 es más corta cuando se mide inmediatamente luego del ejercicio previo. Presumiblemente, esto se debe a los mecanismos que permiten el transporte de oxígeno y su utilización en los músculos activos. En contraste, la τVO2 es más prolongada con el desacondicionamiento físico lo cual puede deberse a un funcionamiento sub-óptimo de los mecanismos que permiten el transporte y la utilización de oxígeno en los músculos activos.
Suscribirse a:
Entradas (Atom)